Entering the Pathway
Several non-carbohydrate carbon substrates can enter the gluconeogenesis pathway. One common substrate is lactic acid, formed during anaerobic respiration in skeletal muscle. Lactate is transported back to the liver where it is converted into pyruvate by the Cori cycle using the enzyme lactate dehydrogenase. Pyruvate, the first designated substrate of the gluconeogenic pathway, can then be used to generate glucose. All citric acid cycle intermediates, through conversion to oxaloacetate, amino acids other than lysine or leucine, and glycerol can also function as substrates for gluconeogenesis. Amino acids must have their amino group removed by transamination or deamination before entering the cycle directly (as pyruvate or oxaloacetate), or indirectly via the citric acid cycle.
Fatty acids cannot be converted into glucose in animals, the exception being oddchain fatty acids which yield propionyl CoA, a precursor forsuccinyl CoA. In plants, specifically in seedlings, the glyoxylate cycle can be used to convert fatty acids (acetate) into the primary carbon source of the organism. The glyoxylate cycle produces four-carbon dicarboxylic acids that can enter gluconeogenesis. Glycerol, which is a part of alltriacylglycerols, can also be used in gluconeogenesis. In organisms in which glycerol is derived from glucose (e.g., humans and other mammals), glycerol is sometimes not considered a true gluconeogenic substrate, as it cannot be used to generate new glucose.
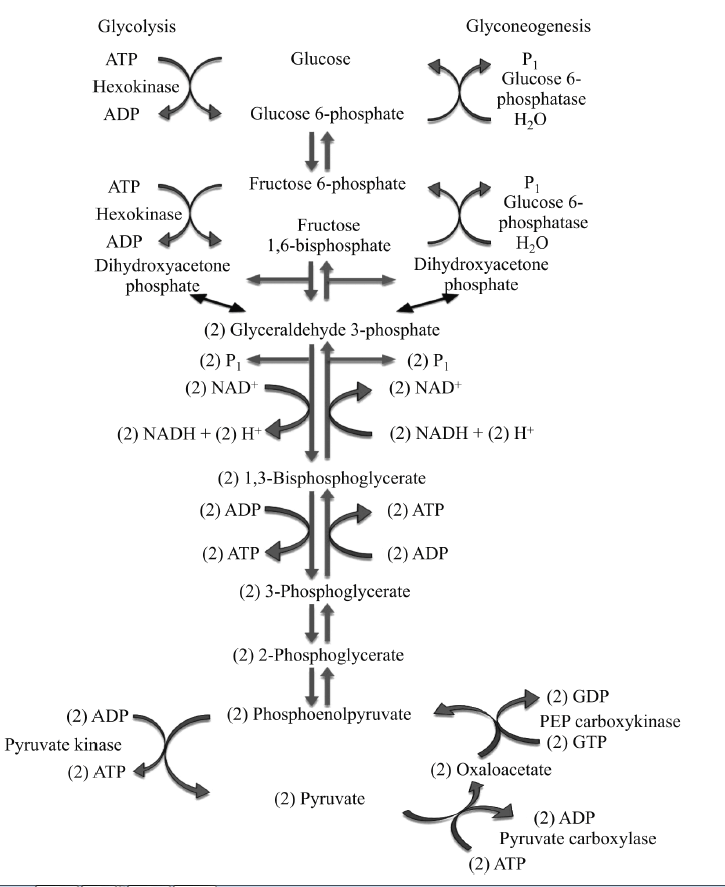
Gluconeogenesis is a pathway consisting of eleven enzyme-catalyzed reactions. The pathway can begin in the mitochondria or cytoplasm, depending on the substrate being used. Many of the reactions are reversible steps found in glycolysis.
Gluconeogenesis begins in the mitochondria with the formation of oxaloacetate through carboxylation of pyruvate at the expense of one molecule of ATP. This reaction is catalyzed by pyruvate carboxylase, which is stimulated by high levels of acetyl-CoA(when fatty acid oxidation is high in the liver) and inhibited by high levels of ADP.
Oxaloacetate must then be reduced into malate using NADH in order to be transported out of the mitochondria.
In the cytoplasm, malate is oxidized to oxaloacetate using NAD+, where the remaining steps of gluconeogenesis occur. Oxaloacetate is then decarboxylated and phosphorylated to produce phosphoenolpyruvate by phosphoenolpyruvate carboxykinase. One molecule of GTP is hydrolyzed to GDP in the course of this reaction.
The next steps in the reaction are the same as reversed glycolysis. However, fructose-1,6-bisphosphatase converts fructose-1,6-bisphosphate to fructose-6- phosphate. The purpose of this reaction is to overcome the large negative ΔG.
Glucose-6-phosphate is formed from fructose-6-phosphate by phospho glucoisomerase. Glucose-6-phosphate can then be used for glucose generation or in other metabolic pathways. Free glucose is not generated automatically because glucose, unlike glucose-6-phosphate, tends to freely diffuse out of the cell.
The final reaction of gluconeogenesis, the formation of glucose, is carried out in the lumen of the endoplasmic reticulum. Glucose-6-phosphate is hydrolyzed by glucose-6-phosphatase to produce glucose. Glucose is then shuttled into the cytosol by glucose transporters located in the membrane of the endoplasmic reticulum.
{kind=link}